May’s Fungi Focus: Bluebell Rust (Uromyces muscari – part 2

Rust fruiting bodies are pretty tiny, and I think it is safe to say that the bulk of them look remarkable similar to the naked eye. As many rusts are limited to a particular plant, identifying the host should be the first step when it comes to identification. I used the example of Alexanders in my piece on Blackberry Leaf Rust and I’ll use it again, because they are rife around coastal areas such as where I live in East Kent. The Romans introduced them to Britain, and presumably they introduced Puccinia smyrnii, the Alexanders Rust, along with them. Another commonly found one that is easy one to identify is Hollyhock Rust, or Mallow Rust (Puccinia malvacearum).
So turning back to our new bible Dutch Rust Fungi, if I were to find a rust growing on my garden mint, I would search for the word mentha in the section on host plants and sure enough, we see a Puccinia menthae listed, the Mint Rust - yes, one potential source of frustration is that to learn about rusts, you are going to have to cultivate a basic knowledge of Latin names for plants too!

Where there are Alexanders, there are often Alexanders Rusts
It’s not always that simple, however, in that some plant families can play host to several different rusts: flowers in the Narcissus family might be affected by Coleosporium tussilaginis or Puccinia schroeteri, while grasses can be afflicted with Puccinia coronata, Puccinia sessilis, Puccinia striiformis and Puccinia graminis – and identifying which type is probably no easier than identifying what the grass in question is in the first place. Puccinia sessilis is more commonly found on arum, such as the woodland species Arum maculatum known as cuckoo pint, lords-and-ladies and a variety of other names, hence its common name of Arum Rust, but it affects Ransoms (Allium ursinum).
By the way, as an alternative or backup to Dutch Rust Fungi, you might want to look at another comprehensive Dutch initiative, the online source Plant Parasites of Europe, which also covers blemishes caused by non-fungal agents such as insects. It might save you a lot of time, as I found out after hours scratching my head while peering down the microscope at a discoloured patch on a holly leaf trying to find spores, only to discover its cause was a small fly known as the Holly Leaf Miner (Phytomyza jucunda).

A Mallow Rust (Puccinia malvacearum) on Common Mallow showing telia and teliospore
Of course, nothing is as simple as it seems, and the life cycle of rust fungi is particularly complex, which is what makes them so fascinating and their study potentially so rewarding. First of all, they can produce up to five fruiting bodies that produce different types of spores. I’ll list the types here just for the sake of completeness, and the coding convention, in brackets afterwards used in the literature to identify what particular stage they are at: spermatium come from spermogonium fruitbodies (0); aeciospores from aecium (I); urediniospores from uredinium (II); teliospores from telium (III) and basidiospores from basidium (IV).
Rusts with a full life cycle forming all five spore types are described as macrocyclic, but many rusts have a reduced life cycle in which some of the stages are skipped, with some only forming teliospores, in a single-stage life cycle that is referred to as microcyclic.
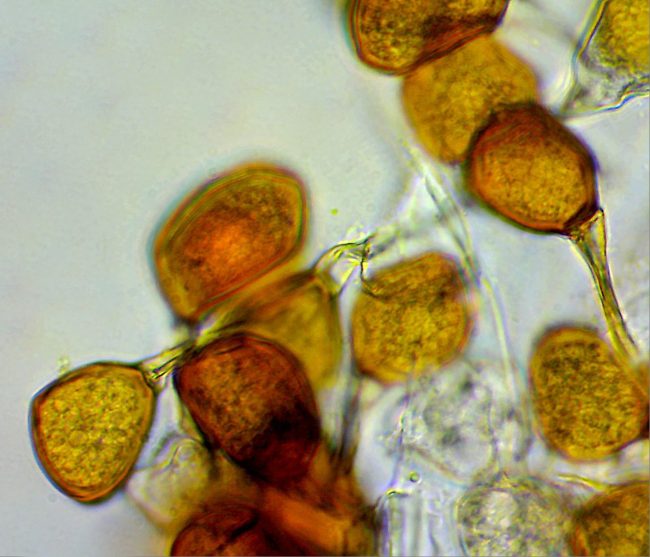
: The Teliospores of the Bluebell Rust are single-celled, typical of rusts in the Uromyces family
The spores all have different forms and functions, and we can largely ignore stages 0 and IV because they are not easily observed by non-specialists. Urediniospores, by contrast are created in the more conspicuous uredinium fruiting body. These only infect other plants of the same species. Teliospores overwinter and then in the spring form basidium fruit bodies which produce basidiospores that can only infect other hosts….
Yes, that’s right, infect other hosts: while some rust fungi, described as autoecious are restricted to a single host species, others jump hosts at a specific point in their life cycle. These are described as heteroecious. We go back to that same Black Rust, Puccinia graminis, that devastated wheat crops for the best example of this. After the telia that form on the dead stems of the wheat (or whatever other grass or serial crop) form their teleospores, the next step, the basidium produce the basiospores that then float off and take up residence on the Common Barberry (Berberis vulgaris). The spermatia formed in this next stage on the barberry make their way back to infect the wheat by way of insects, and so on to the next stage of the cycle. Basically where there is barberry, wheat is more prone to Wheat Rust, so by removing the barberry, you might effectively disrupt the Wheat Rust’s life cycle.

Uredia and urediniospores of the Blackberry Leaf Rust (Phragmidium violaceum)
I’m not going to go into much more detail about these life cycles here, but if the prospect of reading Dutch Rust Fungi is too daunting, there is a rather fun and clearly described account of it all on the Cornell Mushroom Blog by Kathie T. Hodge called ‘Connecting the Rusts’.
I am only mentioning as much as I have here to address one of my own misconceptions and mistakes in last August’s Blackberry Leaf Rust post, specifically relating to its fruit body forms. The purple patches on the leaves are not the individual uredinia but the telia that produce the teliospores - long, segmented, stalked and slightly knobbly. The uredinium fruit bodies of the previous phase of Blackberry Leaf Rust, however, are the characteristic yellowy-orange of many rusts, and the urediniospores are single-celled, spiny, near spherical and but crudely angular.

Telia and teliospores of the Blackberry Leaf Rust Phragmidium violaceum
(I should also mention here something I was not aware of then, that there is another type of Blackberry Leaf Rust, Phragmidium bulbosum, that looks more or less identical until you get the teliospores under the microscope, and notice that rather than 3-5 separate cells, they contain 4-7 segments and the knobbly bit at the tip is a lot longer.)
Urediniospores seem to be a lot less varied, except for in size, than teliospores for most species of rust, and perhaps not much to go on for identifying stuff alone if you need to resort to microscopes, while Termorshuizen and Swertz claim that “Only occasionally the morphology of the aeciospores is valuable for identification” – although in terms of using your naked eyes for identification, the aecium from which they are produced can take a number of forms. For the Puccinia they are typically cup shaped with a wall known as the peridium, while in Phragmidium they don’t have a wall around them at all.

Lords-and-Ladies, or Wild Arum, or Cuckoo Pint, a host for Arum Rust
So bringing this back to a woodland context, a number of rusts associated with woodland plants might be fairly easily found at this point in the year. If the Springtime woodland can be characterised by its brightness and the clarity of its colours, a time when the leaves haven’t really grown back enough from their Autumn shedding to impede direct sunlight from the forest floor, then May is the month when the forest begins to that little bit gloomier and more humid as the tree canopy fills up to signal a new phase in the annual ecological cycle. Those forest flowers that flourished in the sunlight of the first half of the year - Wood Anemones, bluebells and Wild Arum– are now dying back. One assumes that it must be because they have reached the end of their reproductive life and are weakened that the rusts they play host to are now reaching the visible parts of their lifecycles, because by contrast most of the rusts associated with late-blooming garden or wild flowers tend to manifest themselves in late summer.
It would take a particularly perverse type of person to argue that the blotches formed on the leaves of bluebells by the telium of Uromyces muscari are more beautiful than the flowers themselves, but nonetheless, looking at them in close up, macro detail does reveal unexpected colours that have their own aesthetic virtues. Most people probably wouldn’t choose to look that closely anyway, assuming them to be just dry patches on the dying leaves.

Bluebells still form striking carpets of colour on the woodlands floor, but by May they are beginning to look past their best.
Could this be the reason that Dutch Rust Fungi, somewhat surprisingly, lists their distribution in the Netherlands as “not known”? Or that something one might assume was as common as Puccinia asparagi, or Asparagus Rust, is listed as “very rare with the last record in 1937” in the UK? Or that the British Mycological Society’s Lost & Found Fungi Project claims that until 2011, Puccinia cicutae, restricted to the leaves of cowbane, had not been recorded since 1956 and has never been found outside the Fens?
Anyway, we should not fear that the presence of Bluebell Rust is in any way going to jeopardise the beauty of our woodlands for future generations. Left to its own devices, nature ensures its own checks and balances, after all, and we can assume that Uromyces muscari has evolved naturally in synchrony with the life cycle of the bluebell. Rusts may be opportunists, but they clearly have their place in natural ecosystems and we can probably find many examples where the increasing prevalence or scarcity of a particular species is symptomatic of an upset in the balance, and as such, this is undoubtedly an area more of us should take an interest in.

Comments are closed for this post.
Discussion
Thank you thank you for you fabulous article. Nature is such an extraordinarily complex and infinitely fascinating subject. I loved your’lesson’ Receiving monthly nature study pieces like this is what we all need. Indeed I have been mulling over the idea of donating magnifying lenses on chains to a local school. I have been hugely inspired by nature looked at x5! Inspiring more young people to enjoy and care for the environment is essential for its survival.Thing is I don’t want to be responsible for starting fires! What are your thoughts?

Thanks Shuna for your kind words about the article. Sounds like a wonderful idea to give magnifying glasses to school kids. I think the tiny things like this in woodlands and in nature in general are so overlooked and so little understood, we really need to encourage people to look more closely.
I guess every living thing had its role, and everything is dependent on something else – the question is how and why. But at the very least, looking at them more closely, one does get to realise how strangely beautiful it all fits together!
Jasper
12 May, 2020